

OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||

ĆELIJSKA REPRODUKCIJA, ĆELIJSKI CIKLUS
Svi organizmi se sastoje od ćelija koje se umnožavaju deobom. Odrasla
osoba ima oko 100 000 biliona ćelija koje su nastale od jedne jedine oplođene
jajne ćelije. Kod odraslih osoba postoji i veliki broj konstantno dijelećih
ćelija koje zamjenjuju umrle ćelije. Prije nego što se ćelija podijeli
ona mora da se poveća, da udvoji svoj genetski materijal, a potom razdvoji
taj materijal na ćerke ćelije. Ti različiti procesi se odvijaju u ćelijskom
ciklusu. Ćelijski ciklus ili generaciono vreme je najfundementalniji proces
eukariota. On je suština života i smrti
.
Ćelija je osnovna jedinica građe i funkcije svih živih bića. Skup ćelija
sličnog ili istog izgleda, embrionalnog porekla i funkcije naziva se tkivo,
a nauka koja proučava ćeliju naziva secitologija. U ćeliji se nalaze organska
i neorganska jedinjenja. Od neorganskih jedinjenja najzastupljeniji su
voda i soli. Od organskih jedinjenja u ćeliji se nalaze ugljeni hidrati,
masti i proteini. Oba organska jedinjenja sadrže ugljenik. Upoznavanje
građe i funkcije ćelije predstavlja osnovu za svako dublje proučavanje
u biologiji i medicini. Rezultati proučavanja ćelije doprinose poznavanju
i normalnog i patološkog stanja organizma. Kao što je prethodno navedeno
ćelija je osnovna gradivna i funkcionalana jedinica svakog živog bića,
osim virusa. U prirodi postoje brojni organizmi čije se tijelo sastoji
iz jedne ćelije, kao što su bakterije, praživotinje, neke alge i gljive.
Sa druge strane višećelijski organizmi mogu imati više miliona, biliona,
kvadriliona ćelija. Tako se u organizmu odraslog čoveka nalazi oko 10
na 14 ćelija. Iako se sve te ćelije međusobno razlikuju postoje neke osobine
koje su zajedničke svim ćelijama kao što su:
• rast do veličine koja je karakteristična za datu vrstu ćelije;
• obavljanje određenih zadataka (funkcija, uloga);
• primanje signala iz spoljašnje sredine na koje ćelija na određeni način odgovara;
• život ćelije završava se ili ćelijskom deobom ili ćelijskom smrću; pri deobi ćelija daje nove ćelije;
• jedinstven hemijski sastav i jedinstvena građa.
Zahvaljujući razvoju tehnike i instrumenata saznanja o ćeliji su postala veća i potpunija. Tehnika mikroskopiranja je danas dovedena skoro do savršenstva – pronalaskom različitih vrsta mikroskopa (elektronski, fazni, i dr.).
HEMIJSKI SASTAV ĆELIJE
Hemijski elementi koji ulaze u sastav ćelija živih bića nazivaju se biogeni elementi. Od 92 prirodna elementa samo 6 elemenata – C, H, N, О, P i S – ulazi u sastav i čini oko 99% živog tkiva. Prema količini u kojoj su prisutni u ćeliji biogeni elementi se dijele na:
- makroelemente (grč. macro= mnogo) i
- mikroelemente (micro= malo, sitno).
Makroelementi su O, H, C, N, Ca, S, P, K i
dr. Mikroelementi se nalaze u znatno manjim količinama od makroelemenata,
ali je njihovo prisustvo u živim bićima neophodno za normalno odvijanje
životnih procesa. Takvi su npr. Cu, I, Br, Mn, F, Fe i dr.
Oko dvije trećine, odnosno, oko 60% težine odraslog čovjeka čini voda
(kod embriona oko 80%), dok bjelančevine čine oko 17%, masti oko 10%,
ugljeni hidrati oko 1-2% i mineralne materije oko 5%.
Voda
Voda predstavlja najrasprostranjenije jedinjenje u organizmima i neophodan
uslov za njihov opstanak. Voda je jedna od glavnih komponenti živih sistema
i čini čak 50-95% težine ćelije. Osim u samoj ćeliji, voda se nalazi u
međućelijskim prostorima i krvi životinja. U tijelu nekih nižih beskičmenjaka
nalazimo preko 90% vode (dupljari, hidra npr.). Kod mladih listova, stabala
i korjenova voda čini 80-90% svježe mase, a kod sočnih plodova (krastavaca,
lubenice, paradajza) čak preko 90%. Sjemena sadrže svega oko 10% vode,
a ponekad samo 5% (sjeme kikirikija). Količina vode u ćelijama čovjeka
zavisi od: starosti (sa starošću ćelija opada i količina vode u njima);
vrste tkiva (krvno tkivo ima veću količinu vode od npr. masnog tkiva),
metaboličke aktivnosti ćelije (aktivnije ćelije imaju više vode), pola
(žene imaju manje vode od muškaraca).
Da bi organizam čovjeka ispravno funkcionisao potrebno mu je oko 10l vode
dnevno. Dva litra dobija spolja: unese hranom i pićem, dok ostatak stvaraju
sama tkiva. Voda koja nastaje u unutrašnjosti organizma pri kataboličkim
procesima (procesi razgradnje složenih jedinjenja) naziva se endogena
voda (lat. endo = unutra) ili metabolička voda. Sve životinje i biljke
žive od vode koju uglavnom same stvaraju. Endogena voda se zatim razlaže
u tkivima i koristi u različite svrhe.
Neorganske soli
Neorganske soli su takođe veoma zastupljene u ćelijama, a njihovi katjoni i anjoni su neophodni za: održavanje bioloških struktura (gradivna uloga) i biološku aktivnost jedinjenja (metabolička uloga). Najzastupljeniji katjoni su: К+, Na+, Ca++. Među anjonima su to: hloridi, karbonati, bikarbonati i fosfati.
• Na+ i К+ obezbeđuju polarizovanost membrane nervnih i mišićnih ćelija, a time i njihov normalan rad.
• Među anjonima najvažniji su fosfati jer predstavljaju osnovne oblike iz kojih se koristi energija - izgrađuju ATP (adenozintrifosfat).
• Karbonati i bikarbonati imaju ulogu pufera, odnosno, regulišu stalnost pH vrijednost vodenog rastvora. (Pri padu pH vrednosti ispod 7 čovek može da živi samo nekoliko minuta.)
Uloge mineralnih materija
Mineralne materije organizam ne stvara sam, već ih unosi hranom. Radi razumjevanja značaja ovih materija biće navedene uloge nekih najbitnijih:
• Fe (gvožđe) je veoma važan sastojak hemoglobina; nedostatak gvožđa u organizmu ometa normalno stvaranje crvenih krvnih zrnaca, što prouzrokuje malokrvnost – anemiju (mada se mora napomenuti da za ovu bolest postoje i drugi uzroci);
• Ca i P grade kalcijum-fosfate koji su glavni sastojci kostiju;
• S ulazi u sastav nekih aminokiselina;
• Na, К i Cl učestvuju u osmoregulaciji:
• F sprečava karijes zuba;
• Co je sastavni deo vitamina B12 itd.
Organska jedinjenja
Organska jedinjenja obavezno sadrže ugljenik (C) i njihovim razlaganjem
se oslobađa manja ili veća količina energije (razlika u odnosu na neorganske
materije).
Razlikuju se četiri grupe ovih jedinjenja:
• ugljeni hidrati
• lipidi
• proteini
• nukleinske kiseline
METABOLIZAM ĆELIJE
U živoj ćeliji se neprekidno odvija ogroman broj hemijskih reakcija. Cjelina svih hemijskih procesa, odnosno, ukupan promet materije i za materiju vezane energije naziva se metabolizam. Metabolizam karakterišu dva osnovna procesa:
- anabolizam i
- katabolizam.
Anabolizam predstavlja sintezu složenih jedinjenja iz prostih, uz potrošnju
energije kakvi su npr.fotosinteza, sinteza proteina itd. a katabolizam
su reakcije razgradnje složenih jedinjenja na prosta, uz oslobađanje energije,
pripadaju mu procesi kao što su disanje, varenje i dr. U ćeliji se neprekidno
odvijaju tijesno povezani procesi razlaganja organske materije uz oslobađanje
energije i sinteza složenih sastojaka ćelije uz utrošak energije. Pošto
se anabolizam neprekidno odvija (ćelija neprekidno sintetiše proteine,
šećere, masti idr.) ćelija ima stalnu potrebu za energijom. Živa ćelija,
bez obzira na vrstu organizma, energiju dobija oksidacijom organskih jedinjenja,
tj. njihovim sagorjevanjem (što pripada kataboličkim procesima). Organska
jedinjenja se polako i postupno oksidišu tako da se energija iz njih otpušta
sporo, djelimično u vidu toplote, a dijelom i kao hemijska energija (ATP)
koju ćelija može da koristi u anabolizmu.
Po načinu dobijanja organskih molekula, koji služe kao izvor energije
živa bića se dijele u dve velike grupe:
• autotrofe i
• heterotrofe.
Autotrofi su sposobni da vrše fotosintezu (ili hemosintezu), da sunčevu
energiju (ili hemijsku energiju) iskoriste za sintezu organskih materija
koje će im služiti za dobijanje energije.
Heterotrofi uzimaju gotove organske materije hranom i sagorevanjem tih
materija obezbeđuju potrebnu energiju. Hrana heterotrofa direktno ili
indirektno potiče iz organskih materija
REPRODUKCIJA ĆELIJA
Reprodukcija je, po definiciji iz udžbenika "Biologija
za humanom genetikom", fundamentalna karakterstika života koja predstavlja
širenje žive materije u prostoru i vremenu. Osnovni značaj reprodukcije
je prenos genetske informacije sa roditelja na potomstvo. Međutim, ovde
nije riječ samo o organizmima već i o ćelijama. Dakle, da bi ćelija
normalno funkcionisala ona mora da sadrži genetski materijal
istovjetan ćeliji od koje je nastala. Prilikom diobe ćelija tzv. majka
(matična) ćelija dijeli se na dvije ćerke ćelije. Novi organizam, nastaje
spajanjem germinativnih ćelija. Osnovne odlike germinativnih ćelija jeste
sposobnost formiranja novog života i dvostruko manji broj hromozoma,
tj, hromozomskog seta. Ali, kakav je to set? Hromozomski set je
predstavljen određenim brojem homologih hromozoma karakterističnim
za svaku vrstu. Homologi hromozomi su parovi hromozoma koji nose genske
lokuse za iste osobine, a porijeklom su od roditelja (jedan od majke,
drugi od oca). Cjelokupni genetski materijal sisara nalazi
se u nukleo plazmi jedra, u vidu hromozomskog seta. Svaka ćelija organizma
(izuzev spermatocita i oogonija koje imaju polovinu seta) sadrži cjelokupan
hromozomski set identičan u svakoj ćeliji. Pošto se organizam stalno obnavlja,
potrebno je da se ćelije stalno stvaraju (razmnožavaju). Jedino ćelije
mozga i mišića ne podliježu mitozi, odnosno prostoj ćelijskoj diobi. Ali,
prije same diobe neophodno je da se hromozomski set udvoji kako bi novonastale
ćelije imale identičan hromozomski set. Proces udvajanja (replikacije)
DNK molekula odigrava se u interfazi podfaza S. Ova faza se naziva i ćelijsko
mirovanje i predstavlja prvu i najznačajniju fazu mitoze. Nakon interfaze
dolazi do diobe koja se odvija u četiri faze: profaza, metafaza, anafaza
i telofaza .
Kao što je već ptrhotdno navedeno reprodukcija je osnova nasljeđivanja,
bez ovoga se ne bi uopšte moglo pričati o opstanku jedinki, grupa jedinki,
vrsta itd. To je zapravo prenos genetičke informacije od diobe do deobe
ćelije. Tu postoje 2 važna termina:
- MITOZA
- MEJOZA
Ovo su procesi koji su neophodni za prenos nasljedne informacije. Postoje 2 načina prenosa informacije tj. reprodukcije:
- Reprodukcija somatskih ćelija – mitozom
- Reprodukcija germinativnih ćelija – mejozom
MITOZA
Mitoza se označava i kao M-faza, a predstavlja dio ćelijskog ciklusa
u kojem prestaju sintetički procesi u ćeliji i vrši se dioba ćelije. Ona
se odvija veoma brzo i u prosjeku traje 1 - 2 sata. U normalnom ćelijskom
ciklusu iz faze rasta ćelija neminovno prelazi u fazu ćelijske diobe,
koja podrazumjeva kako diobu jedra, njenog genetskog materijala, tako
i diobu citoplazme. Dioba koja se odigrava u somatskim ćelijama naziva
se mitoza. U ovoj diobi od roditeljske
ćelije nastaće dvije ćerke ćelije od kojih svaka ima istu količinu i isti
DNK kao i ćelija od koje su nastale, te će one biti identične roditeljskoj
u svakom pogledu. Zahvaljujući mitozi nastaju nove ćelije neophodne za
rast višećelijskog organizma kao i za obnovu oštećenih tkiva. Međutim,
dioba koja se odigrava u polnim ćelijama u procesu gametogeneze i koja
se naziva mejoza ima za rezultat stvaranje polnih ćelija koje
nisu genetski identične ćeliji od koje su nastale i imaju za polovinu
redukovan broj hromozoma kao i duplo manju količinu DNK. Mitoza je mehanizam
ćelijske diobe koji podrazumjeva podjelu jedra (kariokinezu) i ćelijske
citoplazme (citokinezu). Pri ovoj diobi vrši se distribucija kako genetskog
materijala tako i ostalog sadržaja na dva jednaka dijela, na dvije identične
ćerke ćelije.

Mitoza se odvija brzo i kontinuirano kroz faze koje su označene kao:
- profaza
- prometafaza
- metafaza
- anafaza i
- telofaza
Slika 1 Mitoza

Među njima profaza traje najduže, a metafaza najkraće. Na samom početku
mitoze hromozomi se uočavaju kao dugački konci, po čemu je i sama deoba
dobila ime (grc. mitos = konac)
Slika 2 Ćelija prije ulaska u proces mitoze
Profaza
Slika 3 Profaza

Profaza se karakteriše prisustvom
maksimalno izduženih hromozoma, končastog izgleda mrežoliko isprepletanih
po cijelom jedru
. Na svaki kraj ćelije tj. na polove, odlazi po jeda par centriola
budući da je došlo do njihove duplikacije u G2 fazi interfaze. Migracija
centriola na suprotne polove ćelije odvija se na taj način što parovi
centriola, koji nakon duplikacije stoje u neposrednoj blizini, intenzivno
sintetišu mikrotubule koje se umeću između njih kao lučno uobličene niti
. Kako
Polimerizacija mikrotubula napreduje
ove niti se sve više izdužuju i potiskuju centriole na suprotne krajeve
ćelije. Figura koja u ovom procesu nastaje naziva se mitotsko (diobno)
vreteno. Krajem ove faze hromozomi postaju vidljivi, kraći, deblji i tamnije
su boje, a uzdužno su podjeljeni na dvije identične hromatide koje su
spojene centromerom. Na svakoj centromeri dolazi do formiranja dva kinetohora,
po jedan na svakoj hromatidi, smještenim na suprotnim stranama hromozoma.
Kinetohor predstavlja proteinski kompleks za koji će se vezati određeni
tip mikrotubula, odnosno niti diobnog vretena tzv. kinetohorne mikrotubule.
Ostale mikrotubule predstavljaju polarne mikrotubule i one se
protežu između suprotno postavljenih parova centriola, ali u predjelu
ekvatora ne dolaze u kontakt sa hromozomima. U humanim somatskim ćelijama
za jedan kinetohor se zakači u proseku 30 mikrotubula.
Prometafaza
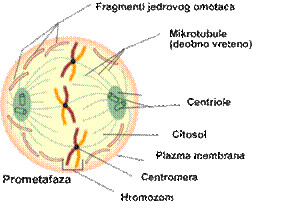 Prometafaza započinje
fragmentacijom jedrovog omotača pri čemu se sadržaj jedra meša sa citoplazmom,
a zatim iščezava jedarce i definitivno se formira deobno vreteno. Hromozomi
vezani za kinetohorne mikrotubule počinju grupisanje u ekvatorijalnoj
zoni.
Prometafaza započinje
fragmentacijom jedrovog omotača pri čemu se sadržaj jedra meša sa citoplazmom,
a zatim iščezava jedarce i definitivno se formira deobno vreteno. Hromozomi
vezani za kinetohorne mikrotubule počinju grupisanje u ekvatorijalnoj
zoni.
Slika 4 Prometafaza
Metafaza
Slika 5 Metafaza

Metafaza traje veoma kratko
i u njoj su hromozomi već u velikoj mjeri kondenzovani tako da su vrlo
dobro vidljivi zbog čega su upravo ovde najpogodniji za analizu. Budući
da su oslobođeni jedrove membrane, oni se, potiskivani od strane kinetohornih
mikrotubula, smještaju u ekvatorijalni (središnji) region ćelije. Tu se
postavljaju jedan pored drugog tako da su im hromatide paralelne ekvatoru
diobnog vretena, pri čemu se formira figura označena kao metafazna
ploča. Hromatide na
Hromozomu se razdvajaju i ostaju
spojene još samo na nivou centromere.
Anafaza
Slika 6 Anafaza

 Anafaza je
takođe kratka i traje svega nekoliko minuta. Na njenom početku centromere
se dijele, što omogućuje i hromozomima da se podijele na dvije hromatide
koje će postati sestrinski hromozomi. Niti diobnog vretena
se tokom ove faze postepeno skraćuju i sestrinske hromozome povlače na
suprotne polove diobnog vretena. Za kretanje hromatida ka polovima potroši
se nekoliko molekula ATP-a. Od svakog hromozoma jedna hromatida odlazi
na jedan, a druga na drugi pol ćelije. Rezultat ovoga je da se na oba
ćelijska pola našao ponovo diploidan broj hromozoma. U humanoj ćeliji,
čijih se 46 hromozoma podijelilo na 92 hromatide, po 46 hromatida (novih
hromozoma) se nalazi na svakom polu ćelije. Krajem ove faze počinje i
diobacitoplazme, stvaranjem kontraktionog prstena koji predstavlja
prstenasto suženje koje se obrazuje oko ćelije na nivou ekvatora diobnog
vretena. On nastaje kao rezultat
Anafaza je
takođe kratka i traje svega nekoliko minuta. Na njenom početku centromere
se dijele, što omogućuje i hromozomima da se podijele na dvije hromatide
koje će postati sestrinski hromozomi. Niti diobnog vretena
se tokom ove faze postepeno skraćuju i sestrinske hromozome povlače na
suprotne polove diobnog vretena. Za kretanje hromatida ka polovima potroši
se nekoliko molekula ATP-a. Od svakog hromozoma jedna hromatida odlazi
na jedan, a druga na drugi pol ćelije. Rezultat ovoga je da se na oba
ćelijska pola našao ponovo diploidan broj hromozoma. U humanoj ćeliji,
čijih se 46 hromozoma podijelilo na 92 hromatide, po 46 hromatida (novih
hromozoma) se nalazi na svakom polu ćelije. Krajem ove faze počinje i
diobacitoplazme, stvaranjem kontraktionog prstena koji predstavlja
prstenasto suženje koje se obrazuje oko ćelije na nivou ekvatora diobnog
vretena. On nastaje kao rezultat
Kontrakcije mikrofilamenata, aktina i miozina, vkoji su vezani za ćelijsku
membranu .
Telofaza
Slika 7 Telofaza

Telofaza, završna faza mitoze (grc. thelos = kraj), počinje
u momentu kada hromozomi dostignu polove diobnog vretena. Za vreme ove
finalne faze mitoze dolazi do despiralizacije hromozoma, oni se izdužuju
i postaju slabije vidljivi. Oko hromozoma, na svakom ćelijskom polu, formira
se jedrova membrana i pojavljuje se nukleolus. Tako su formirana dva nova
jedra i time je kariokineza definitivno završena .
Istovremeno dolazi i do ravnomjerne raspodjele organela kao i ostalog
ćelijskog sadržaja u citoplazmi oko novoformiranih jedara.
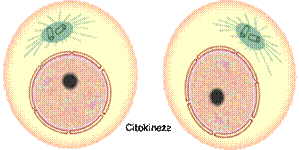
Citokineza koja je započela u anafazi dalje se nastavlja na taj način
što se oko ćelije sve više steže kontraktilni prsten zahvaljujući kontrakciji
aktina i miozina, što uslovljava sve dublju invaginaciju ćelijske membrane.
Podjela citoplazme se završava kada se dvije suprotne strane ćelijske
membrane potpuno približe i spoje, čime su definitivno formirane dvije
diploidne, genetski identične ćerke ćelije. Ćerke
ćelije imaju upola manju količinu citoplazme od majke ćelije. Kada uđu
u interfazu tokom perioda rasta one će dostići veličinu majke ćelije
MEJOZA
Mejoza je ćelijska dioba koja se odvija u polnim ćelija. Obrazovanje
polnih ćelija (gameta) čovjeka naziva se gametogeneza . Razlikujemo
spermatogenezu (obrazovanje spermatozoida) i oogenezu (obrazovanje
jajne ćelije). U procesu mejoze se od diploidnih, matičnih ćelija spermatogonija
i oogonija stvaraju polne ćelije, tj. muške i ženske gamete koje sadrže
haploidan broj hromozoma (n), odnosno za polovinu manje od somatskih.
Značaj mejoze se ogleda u održavanju stalnog broja hromozoma iz generacije
u generaciju. U suprotnom bi se broj hromozoma duplirao u svakoj sledećoj
generaciji, i na kraju desete generacije broj hromozoma kod čovjeka bi
iznosio 23552. Haploidan broj hromozoma nastaje u procesu tzv. redukcione
diobe (lat. reductio = smanjenje) matičnih polnih ćelija. Ova dioba
se odvija kroz dvije brze, uzastopne diobe koje su označene kao prva
i druga mejotička dioba. U ovoj diobi se jedna diploidna
ćelija dva puta dijeli i nastaju četiri haploidne ćelije. Redukcija broja
hromozoma obavlja se u prvoj diobi, označenoj kao mejoza I, kada se majka-ćelija
(2n broj hromozoma) podijeli na dvije ćerke ćelije (n broj hromozoma).
U drugoj diobi, mejozi II, se obje ćerke ćelije (sa n brojem hromozoma)
podijele tako da nastaje ukupno 4 ćelije. Rezultat ove diobe su polne
ćelije - gameti koji su genetski različiti. Još jedan značaj mejoze je
u održavanju ogromne genetičke raznovrsnosti što omogućuje slučajan raspored
majčinih i očevih hromozoma u polnim ćelijama. U ovim hromozomima je prethodno
došlo i do crssing-overa što takođe doprinosi raznovrsnosti. U gametima
čoveka od 23 hromozoma moguće je 223 kombinacija, što zanači da čovek
može da obrazuje 8 000 000 različitih polnih ćelija. 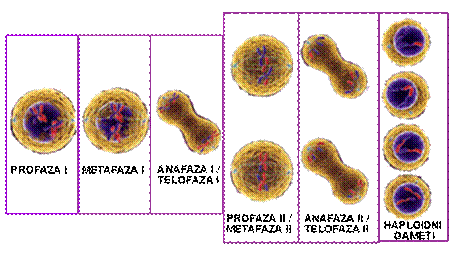
Mejoza I

Prva mejotička dioba se, slično kao i mitoza, sastoji od četiri faze koje su označene kao:
profaza I,
metafaza I,
anafaza I
telofaza I.
Slika 8 Mejoza 1
Ovoj diobi podliježu primarne spermatide i primarne spermatogonije koje su diploidne, a nastale su mitotičkom diobom iz spermatogonija. Mejozi I prethodi interfaza u kojoj je, između ostalog, izvršena replikacija DNK. Svaki hromozom ćelije koja ulazi u mejozu I se sastoji od 2 molekula DNK (dvije hromatide).
Profaza I
Profaza prve mejotičke diobe je duža nego kod mitoze i podjeljena je na pet podfaza:
- leptoten
- zigoten
- pahiten
- diploten
- dijakinezis.
 U
leptotenu počinje kondezovanje hromatina pa se hromozomi uočavaju kao
končaste tvorevine koje su krajevima vezane za jedrovu membranu. Hromozom
se sastoji od 2 hromatide, ali su one priljubljene jedna uz drugu pa se
ne uočavaju. Dakle, u leptotenu se u jedru germinativne ćelije kao i primarne
spermatocite i primarne oocite nalazi diploidan broj uskih i dugačkih
hromozoma koji će se u sledećoj podfazi, grupisati u bivalente.
U
leptotenu počinje kondezovanje hromatina pa se hromozomi uočavaju kao
končaste tvorevine koje su krajevima vezane za jedrovu membranu. Hromozom
se sastoji od 2 hromatide, ali su one priljubljene jedna uz drugu pa se
ne uočavaju. Dakle, u leptotenu se u jedru germinativne ćelije kao i primarne
spermatocite i primarne oocite nalazi diploidan broj uskih i dugačkih
hromozoma koji će se u sledećoj podfazi, grupisati u bivalente.
Slika 9. Faza 1-leptoten
 U
zigotenu dolazi do sparivanja homologih hromozoma tj. grupisanja u bivalente,
homologe hromozomske parove (jedan porijeklom od oca jedan od majke) koji
se čvrsto pripajaju, jednom složenom proteinskom strukturom koja je označena
kao sinaptonemalni kompleks. Proces formiranja bivalenata
je označen kao sinapsis. Budući da je svaki hromozom sastavljen
od dvije hromatide, bivalenti imaju četiri hromatide, zbog čega se nazivaju
i tetrade grc. tetra = četiri). Slika 10. Faza 2 - zigoten
U
zigotenu dolazi do sparivanja homologih hromozoma tj. grupisanja u bivalente,
homologe hromozomske parove (jedan porijeklom od oca jedan od majke) koji
se čvrsto pripajaju, jednom složenom proteinskom strukturom koja je označena
kao sinaptonemalni kompleks. Proces formiranja bivalenata
je označen kao sinapsis. Budući da je svaki hromozom sastavljen
od dvije hromatide, bivalenti imaju četiri hromatide, zbog čega se nazivaju
i tetrade grc. tetra = četiri). Slika 10. Faza 2 - zigoten
 U
pahitenu se između homologih hromatida na određenim mjestima uspostavljaju
veze, hijazme (mostovi), kojima se vezuju homologi genski parovi. Preko
hijazmi se vrši razmjena odgovarajućih fragmenata hromatida između homologih
hromozoma, što se naziva crossing-over, a ima za rezultat stvaranje nove
genske konfiguracije, odnosno genetsku rekombinaciju. Poslije izvršenog
crossing-overa hromozom iz majčine garniture sadrži dio očevog homologog
hromozoma i obrnuto. Slika 11. Faza 3-pahiten
U
pahitenu se između homologih hromatida na određenim mjestima uspostavljaju
veze, hijazme (mostovi), kojima se vezuju homologi genski parovi. Preko
hijazmi se vrši razmjena odgovarajućih fragmenata hromatida između homologih
hromozoma, što se naziva crossing-over, a ima za rezultat stvaranje nove
genske konfiguracije, odnosno genetsku rekombinaciju. Poslije izvršenog
crossing-overa hromozom iz majčine garniture sadrži dio očevog homologog
hromozoma i obrnuto. Slika 11. Faza 3-pahiten

U diplotenu hromozomi se razdvajaju, ali spojevi na hijazmama ostaju.
Slika 12. Faza 4-diploten
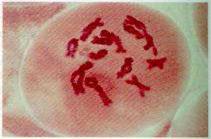 U
dijakinezi hromozomi se maksimalno kondenzuju, a za njih se zakače kinetohorne
mikrotubule i počinje njihovo pomjeranje ka ekvatorijalnom regionu ćelije.
Jedarce nestaje, a jedrova membrana se dezintegriše.
U
dijakinezi hromozomi se maksimalno kondenzuju, a za njih se zakače kinetohorne
mikrotubule i počinje njihovo pomjeranje ka ekvatorijalnom regionu ćelije.
Jedarce nestaje, a jedrova membrana se dezintegriše.
Slika 13. Faza 5-dijakineza
Metafaza I
Metafaza I se karakteriše prisustvom homologih hromozomskih parova spojenih hijazmama, koji se, zakačeni za mikrotubule diobnog vretena, grupišu u područja ekvatora gde formiraju figuru označenu kao metafazna (ekvatorijalna) ploča. Ovde se hromozomi nalaze u parovima od kojih svaki vodi porijeklo od jednog od roditelja- jedan od oca, a jedan od majke. Za razliku od mitoze, gdje su na ekvatoru ćelije bili pojedinačni, u metafazi mejoze I nalaze se parovi homologih hromozoma. Centromere hromozoma su koncima diobnog vretena vezane za polove ćelije i to tako što je jedan hromozom iz para vezan za jedan, a drugi hromozom za drugi pol ćelije.
Anafaza I
Anafaza I počinje razdvajanjem homologih hromozoma, kidanjem spojeva na hijazmama, tako da na svaki ćelijski pol odlazi jedna kompletna, po broju haploidna (n) garnitura hromozoma, ali ona je diploidna (2n) po sadržaju DNK budući da na polove odlaze kompletni hromozomi, a ne hromatide kako je bilo u anafazi mitoze. To znači da u ovoj fazi dolazi do redukcije broja hromozoma. U humanim ćelijama na polovima se nalazi po 23 hromozoma i to iz svakog od 23 para po jedan hromozom.
Telofaza I
Telofaza I počinje i završava se kompletnom citokinezom koja će kod muškog pola rezultovati stvaranjem dvije sekundarne spermatocite od početne primarne spermatocite, a kod ženskog pola od primarne ovocite, nakon ove diobe, nastaje jedna sekundarna ovocita i jedno polarno tijelo. I sekundarna spermatocita i sekundarna oocita, iako haploidne po broju hromozoma, su diploidne po količini DNK, koja će se u narednoj, drugoj mejotičkoj diobi redukovati da bi se dobile definitivne haploidne muške i ženske polne ćelije. U ovoj fazi se formira jedrova opna oko hromozoma koji su na polovima i obrazuje se jedarce. .
 Mejoza
II
Mejoza
II
Nakon telofaze slijedi kratka interfaza u kojoj se, međutim, ne odvija sinteza DNK tako da ova faza ne predstavlja pravu interfazu zbog čega se naziva još i interkineza. Nakon nje sledi druga mejotička dioba koja se, takođe sastoji od četiri faze označene kao:
profaza II,
metafaza II,
anafaza II
telofaza II
Slika 14 Mejoza II

Slika 15. Četiri faze mijotičke diobe
Slika 16. Spermatogeneza
 Pri
ovoj diobi se sekundarne spermatocite i oocite koje sadrže svaka po 23
hromozoma i koje su diploidne po količini DNK, dijele na po dvije ćelije
od kojih će svaka imati po 23 hromozoma, ali duplo manju količinu DNK,
budući da je u anafazi došli do razdvajanja hromozoma na sestrinske hromozome,
čime je količina DNK redikovana na polovinu. Kao rezultat ove diobe kod
muškog pola u procesu spermatogeneze od početne, diploidne spermatogonije
odnosno primarne spermatocite nastaju 4 ćelije-spermatide
Pri
ovoj diobi se sekundarne spermatocite i oocite koje sadrže svaka po 23
hromozoma i koje su diploidne po količini DNK, dijele na po dvije ćelije
od kojih će svaka imati po 23 hromozoma, ali duplo manju količinu DNK,
budući da je u anafazi došli do razdvajanja hromozoma na sestrinske hromozome,
čime je količina DNK redikovana na polovinu. Kao rezultat ove diobe kod
muškog pola u procesu spermatogeneze od početne, diploidne spermatogonije
odnosno primarne spermatocite nastaju 4 ćelije-spermatide
koje imaju haploidan broj hromozom i duplo
manju količinu DNK nego ćelija od koje su nastale.
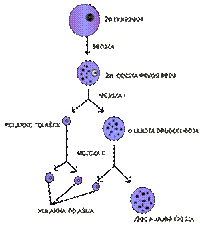 Slično
je i kod ženskog pola kod kojeg u procesu oogeneze od diploidne oogonije,
odnosno primarne oocite nastaje jedna oocita haploidna po broju hromozoma
i količini DNK uz istovremeno obrazovanje tri polarna tela, od kojih dva
nastaju diobom prvog polarnog tijela, a treće deobom sekundarne oocite.
Slično
je i kod ženskog pola kod kojeg u procesu oogeneze od diploidne oogonije,
odnosno primarne oocite nastaje jedna oocita haploidna po broju hromozoma
i količini DNK uz istovremeno obrazovanje tri polarna tela, od kojih dva
nastaju diobom prvog polarnog tijela, a treće deobom sekundarne oocite.
Slika 17. Proces oogeneze
Razlike između spermatogeneze i oogeneze
- Spermatogeneza započinje u pubertetu, kontinuiran je proces, oogeneza započinje za vrijeme embrionalnog razvića, diskontinuiran proces-ima dva stajanja (u diktiotenu profaze I i u metafazi II)
- U oogenezi tokom jednog mjeseca sazrijeva jedna j.ć. U spermatogenezi u toku jednog mjeseca sazrijeva veliki broj spermatozoida
- Na kraju spermatogeneze od 1 primarne spermatocite nastaju 4 funkcionalne haploidne ćelije dok na kraju oogeneze od 1 primarne oocite nastaje 1 funkcionalna ćelija i 3 polocite
- Spermatogeneza traje tokom čitavog života, a ogeneza traje do menopauze (450-500 j.ć. sazri za vreme reproduktivne zrelosti žene) .
ĆELIJSKI CIKLUS
Veličina jedinke zavisi prije svega od ukupnog broja ćelija. Kod odraslog
čovjeka, taj broj približno iznosi oko 100 biliona ćelija, ili u prosjeku
oko 1 milijardu ćelija po gramu tkiva. U svim višećelijskim organizmima
tokom čitavog života odvija se kontinuirano stvaranje novih ćelija i uklanjanje
ostarjelih, a stalnost ukupnog broja ćelija odraslih jediniki predstavlja
rezultat finog balansa između ćelijske proliferacije (mitoze) i ćelijske
smrti (apoptoze). Dioba ćelije obezbjeđuje da novonastale ćerke ćelije
dobiju podjednaku količinu DNK (odnosno hromozoma), a i u većini slučajeva
približno podjednaku količinu citoplazme sa organelama i molekulima. Ćelijski
ciklus obuhvata vrijeme i događaje koji se odvijaju između dvije ćelijske
diobe, uključujući i samu ćelijsku diobu. Njegova dužina zavisi od vrste
i može da traje od 1,5-2 sata kod ćelija kvasca koje rastu, do nekoliko
godina, kod ćelija jetre čovjeka. Međutim, minimalno vrijeme ćelijskog
ciklusa za ćelije čovjeka iznosi oko 24 sata.
Tradicionalno se ćelijski ciklus dijeli u četiri stadijuma ili faze: M,
G1, S i G2. U toku M faze (mitoze), odigravaju se mikroskopom najuočljiviji
događaji, kada se vrši podjela jedrovog materijala – kariokineza, i podjela
ćelije – citokineza. Ovo je ujedno i najkraća faza ciklusa i kod tipične
ćelije sisara traje oko jednog sata. Ostali period između dvije M faze
ćelijskog ciklusa naziva se interfaza. Iako se pomoću mikroskopa ne uočavaju
neke značajnije promjene izuzev uvećanja ćelijske mase, tokom interfaze
se dešavaju mnogobrojni događaji tokom kojih se ćelija priprema za diobu
i sastoji se iz preostale 3 faze ćelijskog ciklusa: G1, S i G2 faze.
Tokom S faze (S=sinteza), odvija se replikacija, odnosno sinteza nukleusne
DNK. G1 faza (G=gap, pukotina engl.) je interval između završene mitoze
(M faze) i početka S faze, dok je G2 faza interval između S faze i početka
M faze. Tokom čitave interfaze, u ćeliji se odvija transkripcija gena
i sinteza proteina neophodnih za uvećanje ćelijske mase, kao i za kontrolu
i odvijanje ćelijskog ciklusa. G1 i G2 faza omogućavaju dodatno vrijeme
ćeliji da uveća svoju masu i udvostruči količinu ćelijskih organela, da
bi posle diobe, ćerke ćelije imale odgovarajuću količinu organela i ćelijsku
masu. (Kada bi interfaza trajala samo onoliko koliko je potrebno ćeliji
da izvrši replikaciju DNK, ćelija ne bi imala vremena da udvostruči svoju
masu, pa bi posle svake diobe ćelije postajale progresivno sve manje i
manje. To se zaista i dešava u posebnim okolnostima. Naime, u toku prvih
nekoliko dioba oplođene jajne ćelije (brazdanje jajne ćelije) dolazi do
progresivnog smanjenja novonastalih ćelija. U toku ovih ćelijskih ciklusa,
G1 i G2 faza su drastično skraćene i praktično ne postoje.)
Ćelijski ciklus je veoma organizovan i složen proces, koji obezbeđuje
kompletnu i preciznu replikaciju DNK i ćelijskih komponenti prije diobe.
U toku ovog procesa učestvuje čitav niz molekula, enzima uključenih u
proizvodnju novih ćelijskih komponenti (DNK, membrane organela, itd.).
Pored ove ''proizvodne'' mašinerije, postoje i molekuli koji učestvuju
u kontroli ćelijskog ciklusa, tzv. kontrolni sistem ćelijskog
ciklusa. Ovaj sistem obezbeđuje pravilnost u napredovanju ćelijskog ciklusa,
sprečavajući napredovanje ukoliko u određenoj fazi ciklusa nisu stekli
svi neophodni uslovi za prelazak u sledeću fazu. Za jedan višećelijski
organizam veoma je važno da se ćelije dijele kada je to potrebno i u onolikoj
mjeri koliko je to potrebno. Nedovoljno ili prekomjerno dijeljenje ćelija
može da dovede do ozbiljnih poremećaja u normalnom funkcionisanju organizma.
Značajan napredak u razumjevanju događaja tokom ćelijskog ciklusa učinjen
je tokom poslednje tri decenije. Početkom '70 godina 20 vijeka, ispitivanjem
sazrijevanja oocita žabe učinjeni su prvi koraci u razumjevanju mehanizama
ćelijskog ciklusa, otkrićem faktora koji indukuju započinjanje mitoze.
Za kasnije otkriće ključnih molekula za odvijanje i kontrolu ćelijskog
ciklusa, Leland Hartwell, Timothy Hunt i Paul Nurse dobili su Nobelovu
nagradu za medicinu i fiziologiju 2001. godine.
Danas znamo da su za odvijanje ćelijskog ciklusa najvažnije tri grupe
proteina i to: ciklinzavisne kinaze (CDK, cyclin dependent kinases), ciklini
i inhibitori ciklin-zavisnih kinaza. Ciklin-zavisne kinaze su enzimi koji
vrše fosforilaciju (dodavanje fosfatnih grupa na specifične aminokiseline
u proteinima) određenih ključnih proteina, pa ih tako najčešće aktiviraju.
Ciklini su proteini koji kada su prisutni u dovoljnom broju aktiviraju
CDK, dok inhibitori ciklin-zavisnih kinaza kada su prisutni u dovoljnoj
koncentraciji sprečavaju njihovu aktivnost
.
Ćelijski ciklus obuhvata promjene koje se odigravaju u ćeliji od momenta
njenog nastanka do njene diobe na dvije nove ćelije. Promjene, koje se
u ćeliji dešavaju za vrijeme diobe, mogu se posmatrati mikroskopom što
sa promjenama tokom interfaze nije slučaj.
Ćelijski ciklus se odvija kroz strogo kontrolisani redosled događanja
koji podrazumjeva rast, odnosno udvostručavanje njenog sadržaja, nakon
čega slijedi ćelijska dioba. Ona rezultira
stvaranjem dvije ćerke ćelije koje su kod svih somatskih ćelija genetski
identične ćeliji od koje su nastale. Novonastale ćelije će pored genetskog
materijala 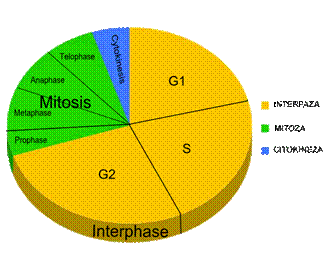 ravnomjerno
podijeliti sve ostale
ravnomjerno
podijeliti sve ostale
komponente roditeljske ćelije kako bi bile sposobne da otpočnu svoj sopstveni
ciklus rasta i diobe. Dakle, novonastale ćelije su identične roditeljskoj
ćelije. Trajanje
ćelijskog ciklusa veoma varira u zavisnosti
od tipa ćelije. Prokariotske ćelije se dijele
amitozom (direktnom ili prostom diobom), dok se eukariotske dijele mitozom
ili mejozom. Kod eukariotskih ćelija koje se brzo dijele, ćelijski ciklus,
traje kratko. Tako ćelijski ciklus kvasca traje svega 90 minuta, dok ćelije
koje se sporo dijele imaju dug ćelijski ciklus i on kod hepatocita (ćelije
jetre) traje oko godinu dana. Međutim, kod većine sisarskih ćelija ovaj
ciklus u proseku traje oko 24 sata.
Slika 18. Ćelijski ciklus
Interfaza
Interfaza je faza između dvije mitoze i u njoj ćelija raste i priprema se za diobu, tako da se biosintetički procesi tokom ćelijskog ciklusa odvijaju samo u ovoj fazi. U životu ćelije ona je najduža faza. Podjeljena je na 3 faze koje su označene kao G1, S i G2.
G1 faza slijedi odmah nakon mitoze i u prosjeku u tjelesnim ćelijama čovjeka traje 9 sati. To je faza u kojoj se se sintetišu mnogi proteini kao i ugljeni hidrati i lipidi neophodni za rast ćelije. Glavna razlika između ćelija koje se dijele sporo i brzo upravo je u dužini trajanja ove faze. Neke ćelije koje se dijele veoma sporo mogu, međutim, napraviti pauzu u svom ćelijskom ciklusu tako da one u G1 fazi ostaju danima, mjesecima, pa čak i godinama. Ova izuzetno duga G1 faza označena je kao G0 faza i u njoj ćelija ostaje sve dok ne bude stimulisana na diobu. Faza G0 se često naziva i faza "mirovanja", što nije najadekvatniji naziv jer zapravo u njoj ćelija nije neaktivna nego naprotiv, ona normalno funkcioniše, vrši sekreciju, fagocitozu, prenos nervnog impulsa itd. Tako su mnogi limfociti u čovjekovom organizmu u fazi G0, ali djelovanjem nekog stimulusa kao što je odgovarajući antigen, oni se mogu vratiti u normalni ćelijski ciklus.
S faza je sintetička faza u kojoj dolazi do sinteze DNK, odnosno duplikacije postojećeg lanca DNK (od jednog molekula nastanu dva potpuno identična molekula DNK), kao i do sinteze histona, nakon čega slijedi organizovanje nukleosoma i obrazovanje hromatinskih vlakana. Replikacijom DNK omogućeno je formiranje dva potpuna genska kompleta, po jedan za svaku novu ćeliju, koja će nastati u procesu mitoze. Ova faza u humanim ćelijama traje oko 10 sati.
G2 faza u većini ćelija traje kratko, a u humanim ćelijama u prosjeku oko 4.5 sata. U njoj se sintetišu proteini neophodni za proces mitoze, kao što je tubulin od kojeg se polimerizuju mikrotubule koje će učestvovati u obrazovanju diobnog vretena. Pored toga, ovdje dolazi do duplikacije centriola kao i drugih organela. Faza G2 i faza G1 obezbeđuju ćeliji vrijeme za rast, sintetičke procese neophodne za stvaranje novih organela i regulatornih proteina, kao i akumulaciju energije. Ako bi interfaza trajala samo toliko vremena koliko je potrebno za duplikaciju DNK, ne bi bilo ćelijskog rasta i pri svakoj narednoj diobi ćelija bi bila sve manja. Međutim, u diobi nekih ćelija upravo se to i događa. Tako su kod blastomera nastalih nakon nekoliko prvih dioba kojima podliježe oplođena jajna ćelija, odnosno zigot, faze G1 i G2 drastično skraćene tako da praktično kod ovih ćelija nema rasta prije dioba, što ima za posljedicu podjelu krupne jajne ćelije u veliki broj sitnijih ćelija, čiji ukupan volumen nije veći od volumena jajne ćelije. Pri kraju G" faze počinje kondenzacija hromozoma, što ujedno označava kraj interfaze i početak mitoze .
-
ZAKLJUČAK
Vjeruje se da su svi organizmi i ćelije koje ih čine nastali od zajedničkog
pretka. Iako su evolutivni procesi nedovoljno poznati i objašnjeni, ipak
se na osnovu podataka koje pružaju fosili i uporednog izučavanje današnjih
ogranizama može pretpostaviti kako je evolucija tekla. Izračunato je da
je Sunčev sistem nastao prije oko 4,6 milijardi godina i poznato je da
od 9 planeta tog sistema život postoji samo na Zemlji (nije isključeno
da na drugim planetama u galaksiji postoji život). Smatra se da je pojavi
života na Zemlji prethodio dug period hemijske evolucije.
Prva ćelija nastala je prije, otprilike, 3,5 ili 4 milijarde godina. Najstariji
do sada otkriveni fosili nađeni su u stjenama starim 3,4 milijarde godina
u Južnoj Africi. Ovi fosili, vidljivi samo pomoću elektronskog mikroskopa,
slični su današnjim prokariotama (bakterije i modrozelene alge).
Na osnovu toga možemo pretpostaviti da je život počeo veoma rano, u prvoj
milijardi Zemljine istorije. Nalazi fosila, takođe, ukazuju da je pre
oko 1,6 milijardi godina došlo do prelaska prokariota ka znatno složenijim
eukariotskim ćelijama. Danas je najprihvatljivija simbiotska teorija koja
objašnjava nastanak eukariotskih ćelija. Po toj teoriji se smatra da su
prokariote ušle u ćeliju eukariota i postale njene organele (mitohondrije
i hloroplasti). Tako je nastala simbioza u kojoj je eukariotska ćelija
obezbjeđivala hranu, a prokariotska energiju. Ova teorija se potvrđuje
građom mitohondrija i hloroplasta koja je slična građi prokariotske ćelije,
ali ima i nedostataka (ne objašnjava npr. pojavu unutrašnjeg ćelijskog
skeleta u eukariotskoj ćeliji). Pored simbiotske postoje i druge
teorije koje pokušavaju da objasne evoluciju prokariotske u eukariotsku
ćeliju. U svakom slučaju, dogod to ne bude moglo da se u eksperimentu
dokaže, biće moguće samo pretpostavljati kako je ovaj proces tekao.
Evolucija ćelija je trajala od 3-4 milijarde godina, dok su se ostali
oblici života, kao i najsavršeniji, razvili u periodu od samo 600 miliona
godina. Izgleda da se evolucija života dugo odvijala u ćeliji, usavršavajući
njenu građu i funkcije, da bi posle toga došlo do stvaranja različitih
organizama za relativno kratko vrijeme.
Što se tiče reprodukcije ćelija, može se reći da je reprodukcija
fundamentalna karakteristika života koja predstavlja širenje
žive materije u prostoru i vremenu te da je snovni značaj reprodukcije
prenos genetske informacije sa roditelja na potomstvo, dok je ćelijski
ciklus život ćelije između dvije diobe, pri čemu je jedna dioba uključena
u ciklus. U gornjem dijelu teksta sam detaljno opisala kako repodukciju
tako i ćelijski ciklus i mislim da u tom dijelu seminarski rad sadrži
veliki broj detalja kako bi se što bolje shvatila suština i način reprodukcije
ćelija i ćelijski ciklus.
LITERATURA
- Milenković Sanja, Stefanović Milica, Cvetković Bora Ćelijski ciklus - srž života Univerzitet u Beogradu, Medicinski fakultet, KBC 'Zemun' Zemun 2007 godine
- Ilustracije iz knjige "Biologija sa humanom genetikom" i na osnovu mikroskopskih preparata Veterinarskog fakulteta u Beogradu)Natura broj 23 mart 2001 godine
- Jelena Grozdanovic-Radovanovic,Citologija, ZUNS, Beograd, 2000 godine
- Milica Matavulj,Ćelija i tkiva (skripta), Novi Sad, 2005 godine
- Prof. dr Vladimir Bumbaširević, Ćelijski ciklus, Institut za Histologiju i embriologiju, preuzeto januar 2011 godine
- Internet, Reprodukcija ćelija, PPT prezentacija, preuzeto januar 2011 godine
- http://www.biologija.rs/mitoza.php
- http://www.biologija.rs/deoba_celije.php