
OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||

DEZOKSIRIBONUKLEINSKE KISELINE - DNK
GENETIKA I NASLJEDNI MATERIJAL
 Genetika
je nauka koja proučava nasljedni materijal,procese i zakonitosti
biološkog nasleđivanja.
Genetika
je nauka koja proučava nasljedni materijal,procese i zakonitosti
biološkog nasleđivanja.
Zahvaljujući nasleđivanju i promjenljivosti javljaju se sličnosti i razlike
osobina u potomstvu u odnosu na roditelje i dalje pretke. Rasvjetljavanje
uzroka tih sličnosti i razlika u potomstvu jeste osnovni cilj genetike
kako bi ona u doglednoj budućnosti mogla da upravlja procesima nasleđivanja
i promjenljivosti.
Pod biološkim nasleđivanjem podrazumjeva se proces razvića novih živih
bića-organizama sličnih roditeljima i daljim precima. Potomstvo liči na
roditelje zbog sličnosti materijalnih osnova nasleđivanja u polnim ćelijama,koje
učestvuju u procesu oplođenja i zato što se u toku razvića jedinke dijelom
ponavlja istorijski razvoj vrste. Razlike između roditelja i potomaka
mogu da budu uslovljene nasljednom promjenljivošću(novim kombinacijama
postojećih nasljednih činilaca i promjenama u nasljednoj supstanci) i
sredinom koja se neprekidno mijenja u toku razvića jednike. Biološko nasleđivanje
i promjenljivost tijesno su povezani sa procesima razmnožavanja i razvića
jedinke.
Nasljeđivanje i promjenljivost su dvije osnovne katakteristike živih bića,međusobno
suprotne ,ali nerazdvojno povezane. Pošto jedinke stare i umiru, priroda
obezbjeđuje neprekidnost života putem razmnožavanja i prenošenja nasljednih
činilaca na nove jedinke-potomstvo.
U nasljedni materijal spadaju: hromatin, hromozom,
DNK i gen.
HROMATIN
Hromatin je supstanca jedra koja sadrži dezoksiribonukleinske kiseline
i proteine.
Tačnije u hromatinu je došlo do uvijanja DNK oko proteina, zato ga jos
i zovemo nukleoprotein. Hromatin se uočava u interfaznom jedru (to je
jedro ćelije koja nije u deobi, već se nalazi u interfazi). On je u obliku
hromatinskih vlakana koja kondenzovanjem (u toku ćelijske diobe) postaju
samostalna telašca – hromozomi.
HROMOZOMI
Proučavajući ćelijsku diobu, naučnik Štrasburger je 1875.godine
zapazio končaste tvorevine koje lako primaju boju,pa ih je nazvao hromozomi
(grčki chroma = boja, soma = tijelo).
Hromozomi čine sastavni dio jedra. Vidljivi su pod mikroskopom obično
u toku diobe ćelije. Hromozomi su izgrađeni od dezoksiribonukleinskih
kiselina, ribonukleinskih kiselina i bjelančevina. Nasljedni genetički
materijal hromozoma sačinjavaju dezoksiribonukleinsle
kiseline.
Hromozomi mogu da se reprodukuju,pa pri diobi jedra osiguravaju neprekidnost
prenošenja nasljednih (genetičkih) informacija iz generacije u generaciju.
Hromozomi imaju vodeću ulogu u procesima nasljeđvanja i promjenljivosti.
Oblik, veličina, građa i broj hromozoma stalni su i karakteristični za
određenu vrstu i poznati pod imenom kariotip (formula vrste).
Hromozomi se po veličini jako razlikuju u zavisnosti od vrste organizma.
Unutrašnja građa hromozoma je složena i najbolje se može uočiti na početku
ćelijske diobe. Tada se svaki hromozom sastoji iz dva kćerinska vlakna
(hromatide) koja su povezana centromerom. U zavisnosti od položaja centromere,tipovi
hromozoma su: metacentričan, submetacentričan, subtelocentričan i telocentričan.
Kod biljaka i životinja koje se polno razmnožavaju različit je broj hromozoma
u tjelesnim (somatskim) i u polnim ćelijama (gametima). Gameti sadrže
jednostruk-haploidan (n) broj hromozoma, a somatične ćelije dvostruk-diploidan
(2n). Haploidnni hromozomi sačinjavaju haploidnu garnituru ili genom.
Broj tjelesnih hromozoma kod čovjeka je 46 a broj polnih hromozoma je
23.
GENI
Gen je fizička i funkcionalna jedinica nasleđivanja, koja prenosi nasljednu
poruku iz generacije u generaciju, a čini ga cjelovit dio DNK potreban
za sintezu jednog proteina ili jednog molekula RNK. Geni su nanizani duž
hromozoma. Gen za određeno svojstvo uvijek se nalazi na istom mjestu na
hromozomu koje se naziva genski lokus.
Različiti oblici jednog istog gena nazivaju se genski aleli i na njima
se zasniva polimorfnost svih organizama. Geni su linearno raspoređeni
delovi hromozomske DNK. Njihova veličina (broj nukleotida DNK) i raspored
na hromozomima su stogo određeni. Građa gena je ustvari građa same DNK
i ogleda se u tačno određenom redosledu nukleotida (A, T, C i G).Vrlo
se rijetko primjećuju iznenadne promjene nasljednih faktora. Promjenjeni
gen nasljeđuje se dalje u istoj formi. Promena tog redosleda, manjak ili
višak nukleotida rezultira u promeni funkcije gena i naziva se genska
mutacija (tačkasta mutacija).Takvim mutacijama
objašnjava se mnoštvo nasljedno različitih rasa.
Sveukupnost nasljednih činilaca (tj. gena) koji ulaze u sastav ćelijskih
hromozoma jednog organizma označavamo kao genotip tog organizma,dok skup
gena u njegovim gametima (tj. u jednoj garnituri hromozoma) čini genom.
Količina DNK u genomu eukariota daleko premašuje zbir gena koji kodiraju
sve proteine prisutne u ćelijama. To znači da dio genoma sadrži nizove
nukleotida koji ne nose šifru za sintezu proteina. Segmenti gena koji
sadrže informaciju za sintezu proteina su nazvani egzoni, a nekodirajući
nizovi između njih su introni.
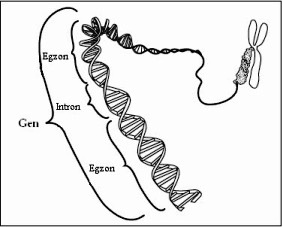
Gen je određen količinom materijala u hromozomu koja se može odvojiti od njega i zamjeniti odgovarajućim dijelom (ali nijednim drugim) homolognog hromozoma.
NUKLEINSKE KISELINE: DNK i RNK
Nukleinske kiseline se danas smatraju
ključnim molekulima života, jer sadrže genetske informacije i neposredno
sudjeluju kod biosinteze proteina.
Nukleinske kiseline se najviše nalaze u jedru (lat. nucleus) pa su po
tome i dobile naziv. Prvi ih je izolovao Fridrih Mišer 1872. godine. Nešto
kasnije ustanovljeno je da se, osim u jedru, nalaze i u citoplazmi. Prema
današnjim podacima poznato je da zasebne nukleinske kiseline sadrže i
neke od ćelijskih organela, kakve su npr. mitohondrije i hloroplasti.
Nukleinske kiseline su krupni i složeni organski molekuli značajni za
ćeliju i odgovorni za najznačajnije procese, kao što su nasleđivanje i
sinteza proteina.
Razlikujemo različite stupnjeve molekulskog reda:
1.primarnu strukturu, tj.sekvencu nukleozida,odnosno
baza; u toj je sekvenci pohranjena informacija koju nosi molekul DNK,
2.sekundarnu strukturu, koja je ovdje definisana kao
način sparivanja baza,
3.tercijarnu strukturu, tj. potpunu prostornu strukturu
sa definisanim položajem svih atoma u prostoru.
Postoje dva tipa nukleinskih kiselina:
— dezoksiribonukleinska kiselina (DNK) i
— ribonukleinska kiselina (RNK).
DNK je nosilac naslednih informacija u ćeliji,
dok RNK učestvuju u prenošenju tih informacija i
njihovom prevođenju u proteine.
Nukleinske kiseline su makromolekuli izgrađeni od
velikog broja građevinskih jedinica koje se nazivaju nekleotidi.
Prema tome nukleinske kiseline su po hemijskom sastavu polinukleotidi.
Nukleotidi su složena jedinjena koja obrazuje jedan pentozni šećer za
koje je vezana fosfatna frupa i jedna azotna,purinska ili pirimidinska
baza. Nukleotidi se međusobno povezuju i na taj način, zahvaljujući vezi
koja se uspostavlja između fosfata i šećera,formirajući lanac.
Osim u virusima, koji sadrže jednu ili drugu nukleinsku kiselinu (nikada
obe), DNK i RNK se nalaze u svim vrstama organizama.
Prema građi su polimeri izgrađeni od monomera - nukleotida.
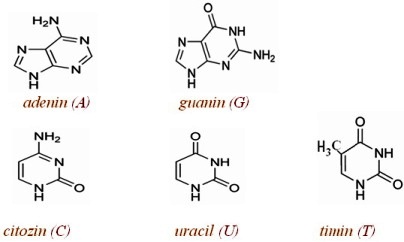
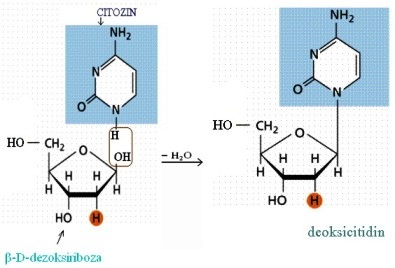
U izgradnji nukleotida, koji formiraju DNK učesvuju:
— pentozni šećer β-D-dezoksiriboza,
— purinske (derivati purina, izgrađene od jednog petočlanog prstena)
baze adenin i guanin ili pririmidinske(derivati pirimidin, izgrađene iz
jednog petočlanog i jednog šestočlanog prstena) baze citozin i timin,
— kiselinski ostatak fosforne kiseline.
U izgradnji nukleotida RNK učesvuju:
— pentozni šećer Driboza,
— purinske (derivati purina, izgrađene od jednog petočlanog prstena)
baze adenin i guanin ili pririmidinske(derivati pirimidina, izgrađene
iz jednog petočlanog i jednog šestočlanog prstena) baze citozin i
uracil,
— kiselinski ostatak fosforne kiseline.

Među heterocikličnim bazama sreću se dvije vrste-derivati purina ili pirimidina.

Hemijski nazivi baza se izvode na osnovu prirode supstituenta i numeracije
atoma osnovnog heterocikličnog jedinjenja,ali se daleko češće koriste
trivijalna imena.
Purinske baze
Najvažnije purinske baze su adenin (6-aminopurin) i guanin (2-amino-6-oksopurin)
koje ulaze u sastav DNK i RNK.
Piramidinske baze
Najvažnije pirimidinske baze su citozin (2-okso-4-aminopirimidin)
,uracil (2,4-dioksopirimidin)i timin (2,4-diosko-5-metilpirimidin). Uracil
se nalazi isključivo u RNK, a timin uglavnom u DNK.
Pored pentoze i baze neophodna je i fosforna grupa da bi nastao jedan
nukleotid;
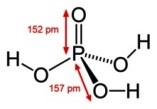
Fosforna kiselina
Međutim, prije nastajanja nukleotida,dolazi do reakcije
jedne baze sa pentozom i dolazi do građenja N-glikozida koji se nazivaju
nukleozidi.
Nukleozidi su prokursori nukleotida i uvijek sadrže β-N-glikozidnu
vezu,dok je pentoza u furanoznom obliku.
Nukleozidi imaju trivijalna imena,koja se izvode iz imena baza;
Kod pirimidinskih nukleozida na korijen imena baze se dodaje sufiks –idin
(npr.cistidin,timidin,deoksicistidin )a kod purinskih nukleozida se dodaje
sufiks –ozin (npr. guanozin,adenozin).
Za razliku od numerisanja prstena baze C-atomi šećera označavaju se sa
1’ do 5’ . Nukleozidi nastaju iz baze i šećera odvajanjem vode i nastajanjem
β-N-glikozidne veze:
Nukleotidi nastaju esterifikacijom jedne
od slobodnih hidroksilnih grupa ostatka pentoze fosfornom kiselinom,pri
čemu mogu nastati 2’-,3’- i 5’ ribonukleotidi,odnosno 3’- i 5’-dezoksiribonukleotidi.
Među njima su najrasprostranjeniji i najznačajniji 5’-nukleotidi (ili
nukleozid 5’-fosfati).
Nomenklatura nukleotida je takva,da na naziv nukleozida dodajemo sufiks
-monofosfat,-difosfat,-trifosfat, u zavisnosti od toga koliko imamo fosfatnih
grupa u nukleozidu.Uobičajno je i skraćeno ispisivanje naziva nukleotida,
npr. dAMP (deoksiadenozinmonofosfat) ili GTP (guanozintrifosfat).

Međutim,primarna hidrokislna grupa pentoze se može esterfikovati i pirofosfornom ili trifosfornom kiselinom,pri čemu se dobijaju difosfati i trifosfati nukleozida. U slučaju adenozina ovim reakcijama nastaju adenozindifosfat (ADP), odnosno adenozintrifosfat (ATP).Nukleotidi se ne nalaze samo u nukleinskim kiselinama.Neki od njih ulaze u sastav tzv. nukleotidnih koenzima,a neki se nalaze slobodni.Tako,na primjer,u svim ćelijama su prisutni,kao stalni sastojci,AMP,ADP i ATP.

Adenozintrifosfat (ATP) je nukleotid poznat u biohemiji
kao „molekulska valuta“ za unutarćelijski transfer energije; to jest,
ATP je u stanju da uskladišti i transportuje energiju unutar ćelija. ATP
takođe igra važnu ulogu u sintezi nukleinskih kiselina. Molekuli ATP-a
se takođe koriste za skladištenje korisne energije koju biljke konvertuju
u ćelijskoj respiraciji.
Nukleinske kiseline su sastavljene od velikog broja nukleotida,povezanih
fosfodiestarskim mostovima preko 3’- i 5’- hidroksilnih grupa pentoza.Prema
tome,skelet polunukleotidnog lanca (kako DNK, tako i RNK) čine naizmjenično
povezani ostaci fosforne kiseline i odgovarajuće pentoze, a azotne baze
su orijentisane bočno u odnosu na skelet ili kičmu lanca.
Kod polinukleotida je karakterisičan 5’-kraj,koji čini prvi nukleotid
i 3’- kraj, koji čini posljednji nukleotid u lancu.

Redoslijed ili sekvenca baze u polinukleotidnim lancima,
od bitnog je značaja za konformaciju nukleinskih kiselina,odnosno za njihovu
biološku funkciju.
Prikazano je povezivanje jednog lanca,tačnije povezivanje nukleotida,ali
DNK je dvostruki,spiralno uvijeni lanac. Svaki polinukleotidni lanac je
pridružen drugome zbog vodoničnih veza koje nastaju među njima i određuju
komplementarno sparivanje prema pravilu: adenin sa timinom;citozin sa
guaninom. Identitet baza pri tome određuje i jačinu i dužinu trajanja
veze.
Zbog sparivanja baza, same baze su okrenute prema unutrašnjosti molekule
tvoreći osovinu zavojnice, a fosfatne grupe i šećeri nukleotida su okrenute
prema van pri čemu lanci čine kostur zavojnice.

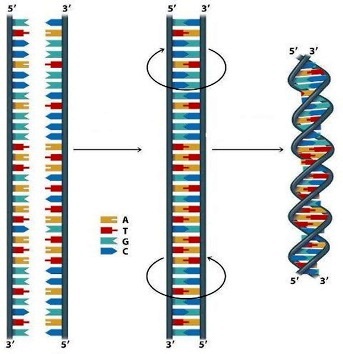
Dakle, DNK se sastoji iz dva polinukleotidna lanca,koji
su uvijeni udesno (u pravcu kretanja kazaljke na satu) oko iste ose.Sparivanjem
baza dobijamo strukturu,gdje su ovi lanci antiparalelni,što znači da njihovi
3’,5’-internukleotidni fosfodiestarski mostovi teku u suprotnim smjerovima.Lanci
nisu identični već su komplementarni.To znači da se nasuprot adeninu u
lancu I,u lancu II nalazi timin,odnosno nasuprot guanozinu-citozin.
Utvrđeno je da se DNK različitih organizama međusobno razlikuju prema
sastavu baza, ali je kod svih ispitivanih DNK ustanovljeno da su molarni
odnosi adenina i timina, odnosno citozina i guanina isti tj. A=T i G=C
. Ovi odnosi se označavaju kao ekvivalencija baza. Interakcija parova
baza dovodi do formacije duplog heliksa.
STRUKTURA, OSOBINE I FUNKCIJA DNK
DNK je materija specifična za svaku živu jedinku na
planeti. Ona je osnovna materija naslijeđa - prenošenja
osobina sa roditelja na potomke.Smještena je unutar ćelijskog jezgra
u strukturnim jedinicama zvanim hromozomi. Na svakom od hromozoma se nalaze
specifični dijelovi koji se nazivaju geni. Svaki gen definiše jednu od
osobina kod čovjeka, koju je naslijedio od svojih roditelja.
Dužina ukupne DNK u jednoj ćeliji se kod čovjeka procijenjuje na 174 cm.
Molekulske težine DNK su vrlo velike i kreću se od 1 do 10 miliona,a u
pojedinačnim slučajevima i od 50 do 100 miliona.Tako DNK Esherichiae coli
ima molekulsku masu 2,4 * 109 i sadrži 8,4 * 106
nukleotida.Najveći molekul DNK nađen je kod vinske mušice, a izgrađen
je od 4 * 1010 nukleotida,dok DNK kod čovjeka sadrži
oko 2 * 1010 nukleotida.
Sama DNK je građena od specifičnih supstanci. Osnovna gradivna jedinica
DNK je nukleotid koji čine dezoksiriboza, fosforna kiselina i jedna heterociklična
baza (adenin,guanin,timin ili citozin) .

DNK je spiralizovana, ima dva lanca, a na svakom od tih lanaca su "nanizani"
nukleotidi, po principu komplementarnosti heterocikličnih baza. Heterocoklične
baze, koje su dio nukleotida, podijeljene su prema svojim hemijskim specifičnostima
na purinske baze i pirimidinske baze. U purinske baze spadaju adenin i
guanin, a u pirimidinske spadaju citozin, timin i uracil (uracil je dio
RNK). Sistem komplementarnosti se zasniva na spajanju jedne purinske sa
jednom pirimidinskom bazom, vezajući tako DNK u dvolančanu spiralnu strukturu.
U dvostrukom lancu DNK se spajaju adenin-timin i guanin-citozin.
Takva jedna specifična sekvenca čini genetički kod, koji će svojim ispoljavanjem
na organizmu dati određenu osobinu.Danas pridajemo veliko značenje metodama
utvrđivanja sekvence DNK,jer njom razjašnjavamo genetski kodon koji se
koristi za pojedine amino kiseline i za utvrđivanje signala koji regulišu,
npr. početak lanca, kraj transkripcije, početak replikacije i dr.
Dakle, gen je jedna precizna sekvenca kombinacija nukleotida koja određuje
određenu osobinu kod organizma. Svaki gen se nalazi na određenom, njemu
specifičnom mjestu na posebnom hromozomu, na DNK. To DNK čini osnovnom
nasljednom makrostrukturom.

Ako se DNK pažljivo izoluju iz biološkog materijala, dobijaju se jako viskozni rastvori, što je u skladu sa njihovom makromolekularnom strukturom. Kada se ovaj rastvor zagrije, ili se dodaju kiseline ili baze, viskozitet opada. Ova činjenica ukazuje da je molekul DNK kruto uređen sistem koji pod uticajem agenasa , koji raskidaju vodonične veze, podliježe promjeni konformacije u nasumice izuvijane polinukleotidne nizove. Ova pojava se naziva denaturacija.
STRUKTURA DNK - KAO MOLEKULSKA OSNOVA ZA OČUVANJE I PRENOŠENJE
GENETSKE INFORMACIJE
Kao što je već rečeno,osnovna uloga DNK je čuvanje i prenošenje genetske
informacije. Ova informacija se nalazi upisana u redoslijedu azotnih baza
i prenosi se na potomke procesom koji se naziva replikacija, odnosno biosinteza
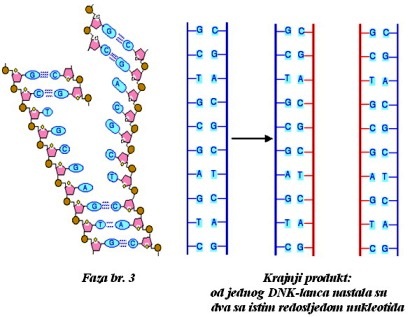
DNK. Replikacija je proces kopiranja, kojim iz roditeljske DNK nastaju
dva idnetična molekula DNK potomka, noseći iste genetske informacije.
Replikacija DNK odvija se prije svake ćelijske diobe i omogućava kasniju
podjelu svakog hromozoma na dvije hromatide. Započinje odmotavanjem lanaca
DNK i njihovim razdvajanjem. Za oslobođene baze u svakom od lanaca vežu
se komplementarne baze koje međusobno poveže DNK polimeraza. Tako na svakom
lancu nastane jedan novi lanac i cijela se DNK udvostruči. Pre nego što
se ćelija podeli, neophodno je da se hromozomi tj. DNK udvostruči (duplira,
kopira) kako bi kćerke ćelije sadržale istu količinu DNK, odnosno sve
kopije gena kao i roditeljska ćelija.
Semikonzervativnost replikacije
Sposobnost DNK da duplira samu sebe od osnovne je važnosti za njenu ulogu nasljednog materijala. Za replikaciju je od najveće važnosti princip komplementarnosti vezivanja naspramnih baza u polinukleotidnim lancima DNK. Udvajanju molekula DNK prethodi rasplitanje dvostruke spirale i odvajanje roditeljskih lanaca. Roditeljski lanci služe kao kalup (matrica) za sintezu novog komplementarnog lanca DNK. Svaki novonastali molekul DNK sadrži po jedan stari i jedan novi lanac DNK, pa se zbog toga kaže da je replikacija semikonzervativan proces (lat. semi= polu; konservativan =očuvan).

Bidirekcionost
Replikacija se i kod prokariota i kod eukariota vrši bidirekciono, što
znači da se od mjesta gde počinje vrši istovremeno u oba smijera i to
isključivo u 5'—» 3' smijeru.
Kod prokariota, čija je DNK prstenasta (kružna) replikacija počinje na
samo jednom mjestu i odvija se bidirekciono. Kada se lanci DNK razmotaju
obrazuje se replikaciona viljuška (ima oblik slova Y).Kod prokariota u
svakom trenutku replikacije postoje dvije replikacione viljuške: jedna
se kreće u smeru kazaljke na satu, a druga u suprotnom smijeru.
Replikacija DNK u eukariotama, čija je DNK linearna, započinje istovremeno
na mnogo mijesta duž hromozoma i teče istovremeno. Na taj način replikacija
ukupne hromozomske DNK se u eukariotskim ćelijama završava za nekoliko
sati, iako je brzina ugrađivanja nukleotida znatno sporija nego kod prokariota.
Enzimi replikacije
Iako je princip replikacije veoma jednostavan, u ćeliji je to složen
proces u kome učestvuje dvadesetak enzima i drugih proteina, od kojih
su najvažniji:
— helikaze,
— nukleaze,
— DNK-polimeraze,
— ligaze i
— primaza.
Helikaze su enzimi koji „hodaju“ po DNK raskidajući
vodonične veze između lanaca DNK u 5’ 3' pravcu.
Nukleaze su enzimi koji raskidaju fosfodiestarske veze,
pri čemu se razlikuju one koje deluju na krajevima lanaca (egzonukleaze)
i one koje deluju na veze unutar lanca (endonukleaze).
DNK- polimeraze su enzimi koji imaju ključnu ulogu u
replikaciji. One u 5’—» 3’ pravcu povezuju nukleotide novog
lanca fosfodiestarskim vezama, pošto su se oni postavili komplementarno
(A-T, C-G) nukleotidima starog (roditeljskog) lanca. DNK polimeraza (kod
prokariota je DNK polimeraza III, a kod eukariota je DNK polimeraza delta)
ima još jednu ulogu: u suprotnom pravcu, 3’ —» 5’, ona raskida
fosfodiestarske veze između pogrešno vezanih nukleotida novog lanca. Ukoliko
DNK polimeraza naiđe na pogrešno sparen nukleotid, ona upotrebi svoju
egzonukleaznu aktivnost u smijeru 3’ —» 5’ i raskine vezu
tog nukleotida sa lancem. Ta njena egzonukleazna aktivnost omogućava ispravljanje
grešaka, koje tokom replikacije nastanu što doprinosi tome da je replikacija
izuzetno tačan proces.
Ligaze deluju suprotno nukleazama – one delove novog
lanca povezuju u cjelinu obrazujući između tih delova fosfodiestarske
veze.
Primaza međusobno povezuje nukleotide RNK u kratke lance
da bi time omogućila dejstvo DNK polimeraze.
Mehanizam replikacije
Endonukleaza zasjeca samo jedan lanac DNK čime počinje njeno rasplitanje.
Helikaze raskidaju vodonične veze između lanaca čime se lanci razdvajaju
i obrazuju se replikativne viljuške. Pošto se replikacija odvija bidirekciono,
na svakom mjestu gde ona počinje obrazuju se dvije replikativne viljuške
koje se kreću u suprotnim smijerovima.
Pratićemo sada šta se dešava u jednoj replikativnoj viljušci. Svaka replikaciona
viljuška je asimetrična jer su lanci antiparalelni, a istovremeno se naspram
oba sintetišu novi lanci u 5’ 3’ pravcu. Lanac DNK koji se sintetiše u
pravcu kretanja replikativne viljuške naziva se vodeći lanac, a onaj koji
se sintetiše u suprotnom smijeru je lanac koji zaostaje. Vodeći lanac
se sintetiše kao cjelovit dok se ovaj drugi sintetiše u vidu dijelova
koji se nazivaju Okazakijevi fragmenti. Sintezu oba lanca obavlja DNK
polimeraza tek pošto se veže za roditeljski lanac koji služi kao matrica.
Ovaj enzim ne može da se veže za ogoljeni lanac-matricu već zahteva postojanje
začetnika (prajmera; engl. primer). Začetnik je kratki lanac RNK i njegovu
sintezu katalizuje enzim primaza. Kada se kratki lanac RNK komplementarno
spari (hibridizuje) sa početkom lanca matrice to omogućuje vezivanje DNK
polimeraze i počinje sinteza novog lanca. Za sintezu lanca koji zaostaje
potrebno je da se sintetiše veći broj začetnika. Okazakijeve fragmente,
po završetku sinteze, međusobno povezuje enzim ligaza.
STRUKTURA I FUNKCIJA RNK
Postoje tri osnovne vrste RNK u ćelijama: ribozomalne RNK (rRNK),
transportne RNK (tRNK) i informacione RNK (iRNK),
koje se nekad nazivaju mesendžer (engleski, message – poruka)
RNK (mRNK). Svaka vrsta ima karakterističan sastav baza,
molekulski masu i biološku funkciju.
Sve RNK su jednolančani molekuli, mada se u nekim dijelovima polinukleotidnog
lanca može obrazovati dvostruki heliks. Udio helikoidalne strukture u
pojedinim vrstama RNK je različit, mada se grubo kreće oko 50%.
Ribozomalne RNK su glavne komponente ribozoma. Njohova
uloga nije sasvim razjašnjena ali se pretpostavlja da sudjeluju u procesu
kretanja ribozoma za vrijeme sinteze proteina.
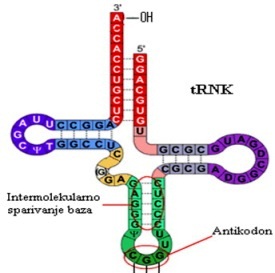
Transportne RNK su relativno mali molekuli koji u molekulu
sadrže od 75 do 90 nukleotidnih ostataka. Ove nukleinske kiseline učestvuju
u procesu aktivacije i prenošenja aminokiselina pri biosintezi proteina.
U organizmima se može naći oko 60 različitih tRNK, pri čemu svaka od dvadesetak
aminokiselina, koje ulaze u sastav proteina, ima barem jednu specifičnu
tRNK, a neke i više. One imaju karakterističnu trodimenzionalnu strukturu
u kojoj je oko 70% u obliku heliksa.
Informacione (mesendžer) RNK se u ćelijama javljaju
u velikom broju, a u strukturi, odnosno sekvenci nukleotida, nose informaciju
za sintezu različitih proteina.
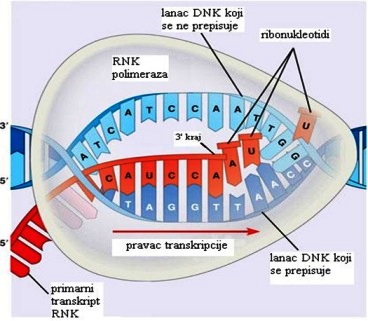
Sve ribonukleinske kiseline se u ćeliji sintetišu procesom koji se naziva
transkripcija, odnosno prepisivanje određenog segmenta DNK . Da bi došlo
do transkripcije, dvostruka spirala DNK se na određenom dijelu odvija
i djelimično razdvaja. Na razdvojenom dijelu, na jednoj niti DNK, koji
je gen za određeni tip RNK, započinje sinteza ribonukleotidnog lanca,
pri čemu se ribonukleotidi ređaju na kalupu po poznatom principu sparivanja
baza ( guanin sa citozinom, a adenin sa uracilom ), a poređani nukleotidi
se povezuju fosfodiestarskim mostovima, zahvaljujući djelovanju enzima
RNK-polimeraze. Nakon završene transkripcije, ribonukleinska kiselina
se odvaja od DNK. Preko pora jedarne membrane prelazi u citoplazmu, gdje,
zavisno od prirode, ispoljava biološku funkciju (izgradnja ribozoma, aktivacija
aminokiselina ili prenos informacije za sintezu proteina).
ZNAČAJ I UPOTREBA DNK
Bioinformatika (en. bioinformatics ili computational
biology) je nauka koja se bavi informatičkim osnovama kao i pohranjivanjem,
organizacijom i analizom bioloških podataka. Na početku su ti podaci dolazili
mahom iz genetike ali se u međuvremenu bioinformatika upotrebljava i u
farmaciji, za proračun prognoze proteinskih struktura i interakcije. Simulacija
i proračun bioloških eksperimenata i podataka se naziva i in siliko proračun
(in silico).
Prvi bioinformatički programi su bili razvijeni za DNK sekvencijalnu analizu.
Tu se radi prvenstveno o brzom pronalaženju ponovljenih dijelova (šablona)
u veoma dugim DNK sekvencama i rješavanje problema preklapanja i pozicioniranja
dvije ili više sekvenci da bi se dobila njihova najveća podudarnost. Primjenu
u toj oblasti su našli algoritmi dinamičnog i metodološkog programiranja.
Kod bioloških hipoteza se rijetko traže tačne podudarnosti kratkih sekvencijalnih
dijelova i to najčešće za odvojene "signale" kao startna i završna
sekvenca genetskog koda.
Genetska identifikacija, genetsko testiranje i genetski
profil su tehnike kojim se vrši identifikacija između individua iste vrste
koristeći pritom samo uzorke njihovih dezoksiribonukleinskih kiselina
(DNK), odnosno genetskog materijala. Ovakva upotreba genetskog materijala
je predstavljena 1985. na Univerzitetu Lajkester od strane njenog izumitelja
gospodina Aleka Džefrisa.
Upotpeba DNK analize
Dvoje ljudi mahom imaju veliku većinu DNK sekvenci zajedničkih. Genetska
identifikacija se služi mikrosatelitima - sekvence koje se ponavljaju
i imaju visok stepen varijacija. Dvoje ljudi koji nisu u krvnom srodstvu
će imati različit broj mikrosatelita na datom lokusu. Koristeći PCR za
utvrđivanje stepena ponavljanja sekvenci na nekoliko lokusa, moguće je
pronaći istu sekvencu koja u normalnim okolnostima nikad ne bi mogla biti
uspostavljena igrom slučaja.
Sudski veštaci koriste u velikoj meri genetsku identifikaciju kako bi
utvrdili moguću vezu između uzoraka krvi, vlakna kose, pljuvačke ili sperme.
Ovaj način identifikacije je takođe pomogao u mnogim slučajevima gde su
pojedinci nepravedno okrivljeni i njihova nevinost ustanovljena. Takođe
se koristi u studijama o populaciji divljih životinja, testiranjima za
utvrđivanje očinstava, identifikaciji leševa, i utvrđivanje sastava prehrambenih
proizvoda. Ova vrsta identifikacija se takođe našla korisnom u postavljanju
hipoteza o modelu ljudske dijaspore u preistorijskom vremenu.
LITERATURA
1. Dr Petar D. Mišić: Genetika,Zavod za udžbenike i nastavna sredstva Srbije,Beograd,1972.
2. Teodosilijus Dobžanski: Evolucija čovječanstva,Nolit,Beograd,1982.
3. Dr Julijana Petrović,Dr Smiljana Velimirović: Hemija za 4. razred gimnazije prirodno-matematičkog i opšteg smjera,Zavod za užbenike i nastavna sredstva Srpsko Sarajevo,2002.
4. www.wikipedia.com
preuzmi
seminarski rad u wordu » » »
